
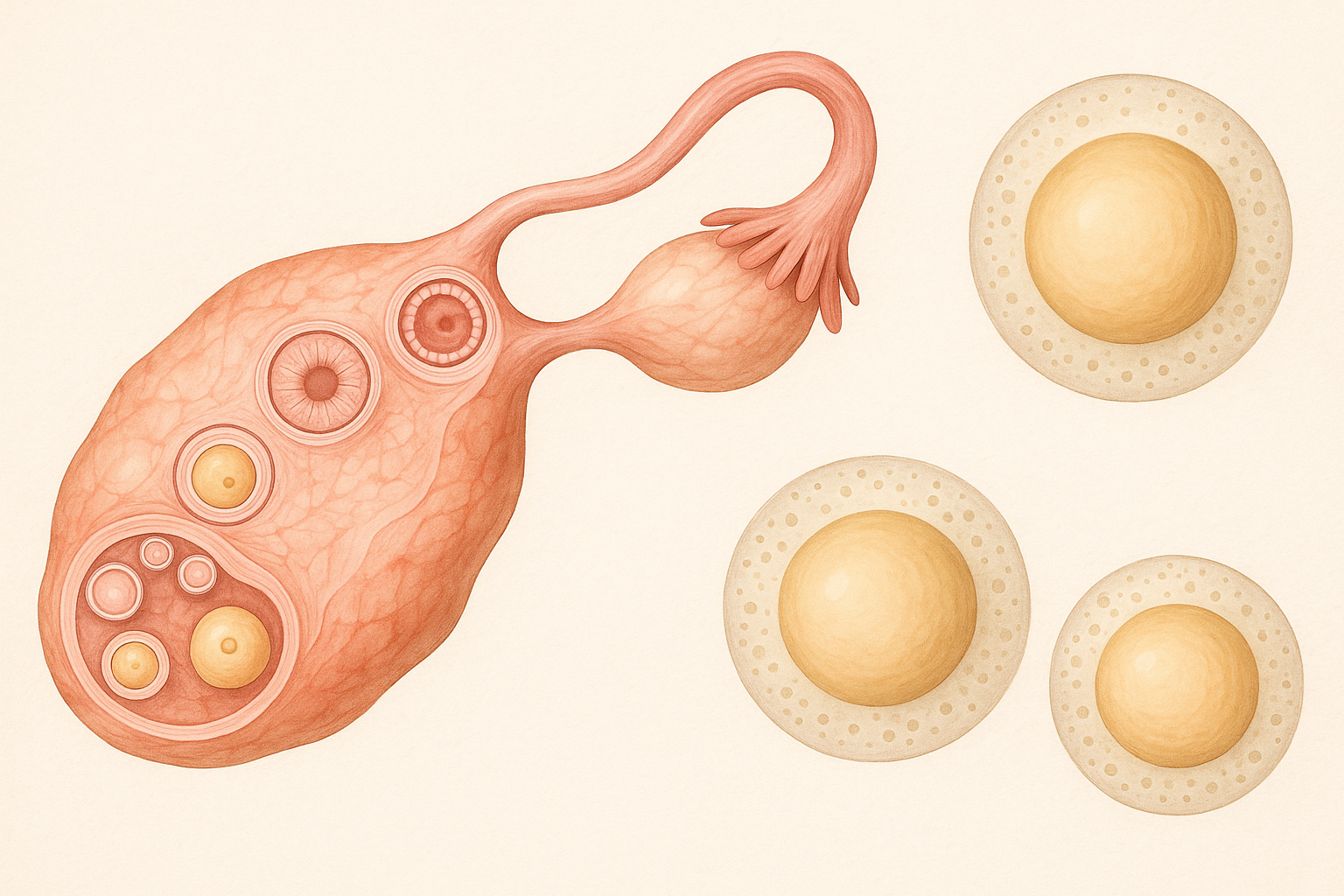
Дозрівання ооцитів – це складний та ретельний фізіологічний процес, який включає два послідовні мейотичні поділи та точно регулюється різними факторами, такими як епігенетика та генетичні фактори. До статевого дозрівання ооцити зупиняються на мейотичній профазі I та містять великий гермінальний візикул, покритий ядерною оболонкою, який відомий як GV ооцит. З настанням сплеску лютеїнізуючого гормону (ЛГ) ооцити зазнають руйнування гермінального візикулу та ініціюють збирання веретена поділу, щоб увійти до стадії метафази I (МІ). Згодом ооцит екструдує перше полярне тільце та зупиняється на стадії метафази II (МІI), завершуючи дозрівання ооцита. Якість та компетентність ооцитів є фундаментальними для визначення подальшої долі ембріона. Дозрівання ооцитів є життєво важливим для досягнення повної компетентності та спричиняє значні зміни в ядрі та цитоплазмі. Цитоплазматичне дозрівання включає зміни в органелах та цитоскелеті, а також інші молекулярні події, що готують ооцити до запліднення та розвитку ембріона.
Синдром порушеного дозрівання ооцитів — це рідкісний стан, при якому яйцеклітини не доходять до зрілої стадії (MII), навіть якщо фолікули ростуть і виглядають нормально під час ЕКЗ.
Генетичні аномалії в дозріванні ооцитів можуть бути результатом різних помилок та генетичних мутацій. Ці дефекти можуть мати глибокі наслідки для ембріонального розвитку. Завдяки застосуванню технології секвенування всього екзому (WES) було виявлено, що генетичні варіації потенційно можуть бути однією з основних причин порушення дозрівання ооцитів. Однак, WES не враховує некодувальні області, епігенетичні зміни, варіанти гонадної мозаїки та недостатньо досліджені гени.
Мітохондрії відіграють життєво важливу роль у виробленні енергії, і будь-які дефекти в їхній функції можуть негативно вплинути на дозрівання та якість ооцитів. У жінок старшого віку оогенез зазнає тривалої зупинки під час профази I, що призводить до накопичення вільних радикалів. Нестабільність мітохондріального геному в поєднанні з тривалим впливом активних форм кисню робить мітохондріальну ДНК (мтДНК) вразливою до різних мутацій. Мутації призводять до збільшення кількості копій мтДНК як компенсаторного механізму; однак частка мутантної мтДНК зазвичай низька порівняно із загальним вмістом мтДНК у зрілих ооцитах, що вказує на низьку гетероплазмію.
Нещодавні досягнення в геноміці та функціональній молекулярній біології виявили групу генів, білкові продукти яких служать важливими молекулярними регуляторами або «охоронцями» компетентності ооцитів.
Зона пелюцида (ZP) являє собою життєво важливу глікопротеїнову матрицю, яка відіграє значну роль у дозріванні ооцитів, овуляції та заплідненні, включаючи індукцію акросомальної реакції, запобігання поліспермії та підтримку ранніх стадій ембріонального розвитку. Зона пелюцида складається з чотирьох глікопротеїнів: ZP1, ZP2, ZP3 та ZP4, кожен з яких кодується специфічними генами. Ці гени ZP експресуються виключно під час розвитку ооцитів протягом росту фолікулів. ZP3 присутній не лише в зоні пелюцида, але й у ядрі ооцитів GV. Крім того, мРНК, що кодують білки ZP, відсутні в ооцитах, що не ростуть, тоді як їх рівень різко зростає в ооцитах середньої стадії та повністю вирощених ооцитах. Дослідження показують, що коли Zp3 пригнічується за допомогою специфічної siRNA, розвиток ооцитів значно порушується, що призводить до помітного зниження відсотка ооцитів MII. Мутації ZP3, можуть значно пошкодити щілинні контакти, що призводить до порушень обміну інформацією між ооцитами та клітинами кумулюс-гранульозу. Це може бути важливою причиною, яка зрештою призводить до зупинки дозрівання ооцитів.
Тубулін бета класу 8 (TUBB8) є одним з генів, які найчастіше відповідають за порушення дозрівання ооцитів. Тубулін (α/β) утворює структурну одиницю мікротрубочок, необхідну для формування веретена поділу під час мейозу. TUBB8 – це ген, що найчастіше піддається мутаціям. Близько 30% зупинок дозрівання спричинені мутаціями в TUBB8, що вказує на домінуючу роль цього гена. Зберігання материнської мРНК та регуляція трансляції під час розвитку ооцитів точно керують експресією TUBB8. Важливо, що аберантний TUBB8 порушує цілісність полюсів веретена та прикріплення кінетохорів до мікротрубочок, тим самим зупиняючи клітину в мейозі I. Мутації TUBB8 на молекулярному рівні порушують складну мережу внутрішньоклітинних шляхів, що сходяться навколо координації мейотичного веретена поділу, динаміки мікротрубочок та цілісності цитоскелета в ооцитах. Мутації з втратою функції в TUBB8, порушують ацетилювання та дестабілізують полюси веретена, що призводить до хромосомного зміщення та зупинки мейозу.
KIF11 належить до родини кінезинів. Він приймає участь як у мітозі так і у мейозі, де він регулює морфогенез веретена поділу ооцитів та вирівнювання хромосом. KIF11 механічно створює сили ковзання назовні, які розділяють центри організації мікротрубочок шляхом зшивання антипаралельних мікротрубочок. Пригнічення або функціональний дефіцит KIF11 призводить до неправильного вирівнювання хромосом у метафазній пластинці, що призводить до помилкових прикріплень між кінетохорами та мікротрубочками веретена, що сприяє анеуплоїдії та блокуванню розвитку ембріонів. Крім того, на активність KIF11 впливають посттрансляційні модифікації тубуліну, які змінюють спорідненість моторного білка до мікротрубочок. Мутації KIF11 опосередковано індукують зупинку мейозу під час фаз GV або MI, зменшуючи активність CDK1(ферментів, який керує клітинним циклом). Мутації KIF11 погіршують генерацію сили та динамічну активність мікротрубочок. Експресія KIF11 зменшується зі старінням ооцитів, що вказує на зв'язок між віковою нестабільністю веретена поділу та динамікою, опосередкованою кінезином. Патогенні мутації в KIF11, погіршують здатність зшивати мікротрубочки та генерувати спрямовану силу, що призводить до статичних, зруйнованих веретен поділу, які не можуть сприяти вирівнюванню хромосом, або до мультиполярних веретен. Саме ці аномалії пов'язані з зупинкою метафази I, неправильно вирівняними кінетохорами та утворенням ооцитів з множинними пронуклеусами (MPN). Зниження якості ооцитів ще більше посилюється зниженням активності KIF11, що пов'язують зі збільшенням рівня активних форм кисню (ROS) та пошкодженням ооплазми.
Мутації в білку 5, асоційованому з цитоскелетом (CKAP5) здатні призвести до значних аномалій форми веретена, невдалої екструзії полярного тільця та, зрештою, мейотичної зупинки — таке явище зазвичай спостерігається вциклах ЕКЗ з незрозумілою зупинкою метафази I. CKAP5, потужна полімераза мікротрубочок та фактор зв'язування, що сприяє динамічному подовженню та стабілізації мікротрубочок веретена завдяки своїй взаємодії з вільними димерами тубуліну. Клінічно, пацієнти з мутаціями KIF11 або CKAP5 демонструють фенотип, що відповідає ранній ембріональній фрагментації або тяжкому блокуванню дозрівання ооцитів.
Мутації CKAP5 дестабілізують зародження мікротрубочок, кероване центром організації мікротрубочок, тоді як дефіцит KIF11 погіршує розвиток біполярного веретена поділу та ацетилювання тубуліну. Кожен з цих компонентів індикує активацію контрольних точок, зупинку мейозу та подальшу некомпетентність розвитку. Будь-яка молекулярна зміна в цій мережі може суттєво вплинути на виживання ооцитів та ембріональний розвиток через специфічну залежність ооцитів від формування ацентросомного веретена та тимчасово регульованої трансляції материнської мРНК. Дані вказують на те, що порушення експресії або функції CKAP5 можуть сприяти деяким порушенням дозрівання ооцитів у пацієнтів з ЕКЗ, у яких відсутні мутації в стандартних генах веретена поділу, таких як TUBB8.
PANX1 (паннексин 1), беруть участь у формуванні єдиних мембранних каналів для молекулярного обміну, включаючи транспорт аденозинтрифосфату (АТФ). Патогенні гомозиготні або складні гетерозиготні варіанти PANX1 призводять до підвищеного вивільнення АТФ у позаклітинний простір, що призводить до дегенерації ооцитів. Ці ооцити часто залишаються незрілими або дегенерують незадовго до або після запліднення.
Ген інтерактора рецептора гормонів щитовидної залози 13 (TRIP13) широко експресується в різних тканинах, включаючи статеві клітини, і відіграє вирішальну роль у мітозі та мейозі. Під час мітозу білок TRIP13 допомагає заглушити контрольні точки веретена поділу, тоді як у мейозі він бере участь у шляхах рекомбінації. Патогенні міссенс-варіанти в першу чергу впливають на мейоз ооцитів, викликаючи незначне зниження рівня білка.
Експресія генетичного білка, асоційованого з топоізомеразою II (PATL2) особливо висока в незрілих ооцитах, та значно знижується у міру дозрівання ооцита. Гомозиготні або складні гетерозиготні варіанти гена PATL2 призводять до зупинки ооцитів на стадії гермінального візикула (GV) або метафази I (MI), невдачу запліднення та ранню зупинку ембріонального розвитку.
ZFP36L2 — це ген, що кодує білок із групи РНК -зв’язувальних білків.РНК-секвенування та GO-аналіз на ооцитах GV пацієнтів з варіантами ZFP36L2 показали, що диференційно експресовані гени задіяні в кількох ключових біологічних процесах, включаючи поділ клітин, сегрегацію хромосом, мітохондріальну функцію та метаболізм рибосом. Більше того, рівень РНК кількох ключових генів, відповідальних за компетентність розвитку ооцитів, був значно знижений. Варіанти ZFP36L2 порушують гомеостаз мРНК в ооцитах та призводять до порушення дозрівання ооцитів.
Епігенетичні модифікації відіграють ключову роль у регуляції експресії генів незалежно від змін у послідовності ДНК, і ці модифікації є невід'ємною частиною дозрівання ооцитів. Метилювання ДНК являє собою первинний епігенетичний механізм, за якого зміни в патернах метилювання можуть порушити експресію генів, необхідну для належного розвитку ооцитів. Аналогічно, модифікації гістонів є важливими, оскільки зміни в посттрансляційних модифікаціях гістонів можуть змінювати архітектуру хроматину та доступність ДНК, тим самим впливаючи на процеси, що є фундаментальними для дозрівання ооцитів.
За оцінками, діагностична ефективність секвенування всього екзому може пояснити від 10% до 40% випадків серед різноманітних генетичних захворювань, що можна використовувати як діагностичну стратегію першої лінії в клінічній практиці.
Для пацієнтів з такою клінічною картиною є варіанти використання дозрівання in vitro (IVM). Але результативність буде нижчою, ніж при застосування стандартних протоколів у пацієтів без порушення дозрівання ооцитів. Як альтернатива для таких пацієнтів може бути також використання донорських яйцеклітин.
Для вирішення порушення дозрівання ооцитів лікар-репродуктолог обирає інитвідуальний підход для кожного окремого клінічного випадку, задля вашого омріяного батьківства.